 science-review.ru
science-review.ru
Scientific journal
Научное обозрение. Медицинские науки
ISSN 2500-0780
ПИ №ФС77-57452
BIOMECHANICS OF PHYSIOLOGICAL LABOR CONTRACTIONS (MYOMETRAL-HEMODYNAMIC CONCEPT)

Наша авторская версия миометрально-гемодинамической концепции биомеханики физиологической родовой схватки в качестве оригинального интеллектуального продукта появилась в информационном поле теории родов у человека в 1980 г. в виде заявки в Госкомизобретений СССР (32ОТ280), а затем, спустя 3 года, и в виде монографии «Биомеханика родовой схватки» (1983). Полная версия концепции была опубликована в 1988 г. в монографии «Биомеханика раскрытия шейки матки в родах» [1]. Случилось так, что именно в это время отечественное акушерское сообщество предложило свой вариант гибридной пейсмекерно-нейрогенной концепции биомеханики физиологической родовой схватки. Этот вариант концепции, фактически без обсуждения и критики, сразу же обрел статус аксиоматического догмата и вот уже пять десятков лет без каких-либо изменений и дополнений позиционируется на страницах всех, в том числе современных учебников и национальных руководств по акушерству [2–4] в качестве основной научной доминанты теории родов. «Гибридность» этой концепции состоит в том, что в ней представлена попытка совмещения одновременного функционирования всех предполагаемо существующих биомеханических феноменов, принимающих участие в процессах дилатации шейки матки, а именно «контракции – ретракции – дистракции», «тройного нисходящего градиента» и «однонаправленной перистальтической волны сокращения» [5–7]. Базовыми условиями, позволяющими реализовать силовые проявления этих биомеханических феноменов, являются представления:
- об особенностях миоархитектоники миометрия рожающей матки человека;
- функциональной гетерогенности различных отделов, слоев и даже различно ориентированных пучков гладкомышечных волокон;
- феномене «ретракции»;
- «реципрокности» различных слоёв и отделов миометрия;
- наличии в миометрии особого рода системы, проводящей электродные потенциалы действия;
- прямой двигательной иннервации уте- ромиоцитов;
– наличия в миометрии водителя рит- ма – пейсмекера.
Оригинальность гибридной концепции [8–10] состоит в том, что фазное родовое сокращение мышечной оболочки рожающей матки человека инициируется, модулируется, синхронизируется и координируется исключительно деятельностью пейсмейкера – водителя ритма и центрами симпатической и парасимпатической вегетативной нервной системы [11]. Недавно мы ещё раз обратили внимание отечественного акушерского сообщества на квазинаучность и абсолютную практическую непригодность доминирующей в учебно-методической литературе гибридной пейсмекерно-нейрогенной теории родов [12]. Сложность ситуации, связанная с появлением в открытом информационном поле теории родов нашей миометрально-гемодинамической концепции биомеханики родовой схватки, состояла ещё и в том, что предложенная нами доктрина по своему содержанию и идеологии была абсолютно альтернативной доминирующей пейсмекерно-нейрогенной теории [11, 13, 14]. Получилось так, что буквально всё, что декларировалось в официальных документах, касающихся биомеханики родовой схватки человека, практически полностью отрицалось и опровергалось в миометрально-гемодинамической концепции, в которой предлагались принципиально иные представления как о генезе родовых сил, так и о механизмах конвертации их энергии в полезную внешнюю работу по «дистракции» шейки матки. Парадоксально, но на всём протяжении своего существования наша концепция не только остаётся в ранге непризнанной, но и официально не обсуждается и не критикуется, несмотря на её альтернативность и довольно широкую презентацию в печати [15–17]. И, наконец, главное, в мировом акушерстве сегодня, в начале третьего десятилетия XXI в., полностью отсутствуют адекватные представления о биомеханике родовой схватки человека, которые практикующий в родильном зале врач мог бы использовать для контроля и регуляции родового процесса. Сказанное, прежде всего, имеет прямое отношение ко всем положениям гибридной пейсмекерно-нейрогенной концепции.
Мы полагаем, что, во-первых, публикация этого обзора может стать триггером для начала обсуждения и критики смыслового содержания самой миометрально-гемодинамической концепции биомеханики родовой схватки, что также возбудит интерес акушерского сообщества к разработке актуальных проблем акушерства в области теории родов человека, без решения которых прогресс в практическом родовспоможении почти невозможен. И, во-вторых, исследования А.Г. Савицкого [15–17] чётко показали, что положения этой концепции даже на существующем этапе разработки уже позволяют предложить ряд достоверных диагностических и лечебных методик, которые можно и нужно использовать в практическом акушерстве.
Мы учитываем существующие форматы журнальных статей, в связи с чем, конечно, изложение представляется в тезисном, но идеологически полноценном варианте. Но все представляемые тезисы имеют никем не оспоренную документальную основу, подробно изложенную в наших монографиях [1, 12, 16].
О функциональной морфологии рожающей матки человека
- До начала родов матка представлена двумя морфофункциональными отделами – плодовместилищем, оболочку которого составляют оболочки тела матки и её нижнего сегмента, и запирательного сегмента, представленного шейкой матки. С началом родов с помощью контракционного кольца плодовместилище делится на два самостоятельно функционирующих отдела – тело матки и её нижний сегмент. Оболочка тела матки и её нижнего сегмента представлена в основном гладкомышечной тканью, в шейке матки организованная гладкомышечная система полностью отсутствует. Таким образом, в начале родов в матке начинают функционировать три самостоятельных функциональных отдела – тело матки, нижний сегмент и шейка матки.
- К началу родов в шейке матки происходят грандиозные по масштабу изменения в её морфоструктуре. Шейка матки из жёстко-упругой субстанции трансформируется в вязко-эластичную тканевую структуру с необходимой для осуществления процесса «дистракции» величиной модуля упругости тканей. Одновременно в толще шейки развивается мощное венозное депо, состоящее из лакунарно трансформированных тонкостенных вен, часть которых имеет прямые артериально-венозные шунты, которые превращают шейку матки в кавернозно-подобную структуру. Доказано, что эффективность процесса «дистракции» шейки матки прямо зависит не только от снижения модуля упругости её тканей, но и от полноты завершения в ней кавернозно-подобных преобразований сосудистой сети.
- Мышечная оболочка тела матки двухслойная. Наружный слой, «рубашка матки», представлен плотно уложенными функциональными сократительными модулями миометрия, верхние точки фиксации которых локализуются в области отхождения от матки круглых связок, а нижние – в толще связок, фиксирующих матку к костному тазу. Между наружным и внутренним мышечными слоями тела матки локализуется гигантское венозное депо – разделительный венозный синус, состоящий из огромных по размерам тонкостенных венозных лакун, армированных тканями функциональных сократительных модулей обоих мышечных слоев, которые многократно переходят из одного слоя в другой. Внутренний мышечный слой, в отличие от наружного, более рыхлый и, наряду с мышечной тканью, содержит в толще огромное количество лакунарно трансформированных вен, превращающих его в губкоподобное образование. Венозные лакуны внутреннего мышечного слоя тела матки широко анастомозируют с венозными лакунами разделительного венозного синуса. Верхние точки фиксации функциональных сократительных модулей, образующих внутренний мышечный слой тела матки, локализованы также в месте отхождения от угла матки круглых связок, а их нижние точки фиксации – в соединительнотканном матриксе шейки матки в области истмико-цервикального соустья. Морфологические точки фиксации оконечностей функциональных сократительных модулей выполняют «якорные функции».
- Мышечная оболочка плодовместилища к началу родов находится в растянутом состоянии. Её функциональные сократительные модули имеют длину в два раза большую, чем величина их длины после полного опорожнения полости плодовместилища от содержащегося в ней концепта.
- Миоархитектоника наружного и внутреннего мышечных слоев тела матки идентично представлена функциональными сократительными модулями, которые от верхних точек анатомической фиксации по спирали распространяются до зоны контракционного кольца и после, участвуя в организации его морфоструктуры, следуют к своим нижним морфологическим точкам якорной фиксации.
- В зависимости от изменения «шага спирали» ход модуля в миометрии обоих слоев в начале косопродольный, затем, по мере приближения к зоне контракционного кольца, он сменяется на косопоперечный и, после входа в состав контракционного конца, становится строго циркулярным. Модули внутреннего мышечного слоя, покинув зону контракционного кольца, образуют оболочку нижнего сегмента матки, и ход их в этой зоне близок к косопоперечному. По своему ходу от одной анатомической точки фиксации к другой модули обоих слоев, перекрещиваясь в плоскости, образуют двухмерную петлисто-ячеистую сеть, которая дополняется трехмерной структурой модулей обоих слоёв, переходящих по мере своего снижения из наружного слоя во внутренний и обратно. Имея в виду переходы сократительных модулей из слоя в слой можно говорить о том, что оболочка тела матки имеет трехмерную структуру, а нижнего сегмента – двухмерную. Такие оболочки, с криволинейной формой и ходом своих структурных элементов, образующие полости, в которых содержится гидравлическое содержимое, предназначены для выполнения функции либо сжатия содержимого полости, либо противостояния давлению, связанному с увеличением объема содержимого этих полостей.
- В миометрии рожающей матки человека нет продольно ориентированных структур, механически связывающих дно и шейку матки. Более того, контракционное кольцо прерывает прямую механическую связь между мышечными структурами тела матки и её нижнего сегмента, становясь для модулей наружного и внутреннего мышечного слоев тела матки нижней функциональной точкой фиксации, а для фрагментов модулей внутреннего мышечного слоя, образующих оболочку нижнего сегмента, наоборот, верхней функциональной точкой фиксации. Гладкомышечные элементы мышечной оболочки рожающей матки человека к началу физиологически протекающих срочных родов полностью денервированы. Все нервные стволы, волокна и окончания, имевшиеся в оболочке небеременной матки, к концу беременности, после последовательно протекающих дистрофически-дегенеративных процессов в них, постепенно «исчезают», подвергаясь лизису.
- В миометрии рожающей матки человека полностью отсутствуют признаки наличия такого морфофункционального образования, как «пейсмекер – водитель ритма».
- Артерио-артериоллярно-капиллярное звено муральной системы кровообращения миометрия рожающей матки человека развито адекватно функциональной массе мышечной ткани. В то же время дренажная муральная система миометрия превращена в гигантское внутримиометральное венозное депо, развитие которого с позиции обеспечения потребностей венозного дренажа и нецелесообразно и избыточно, даже с учетом дренажных потребностей маточно-плацентарного контура кровообращения. С позиции формальной морфологии в рожающей матке человека следует различать наличие трёх контуров венозного дренажа органа. Верхний – отток крови по которому осуществляется по венам яичниковой и подвешивающей связок. Эти вены впадают либо в почечные вены, либо прямо в нижнюю полую вену выше места давления и периодической окклюзии последней задней стенкой рожающей матки. И два нижних, разделенных во время родовых схваток сокращающимся контракционным кольцом. Кровь из этих нижних контуров попадает в тазовые вены, давление в которых намного выше, чем в нижней полой вене.
Биомеханически значимые особенности гемодинамики в контурах кровообращения рожающей матки человека
- В рожающей матке человека различают три основных контура кровообращения – маточно-плацентарный, миометральный и шеечный.
- Интенсивность кровотока в маточно- плацентарном контуре находится под прямым контролем величины внутримиометрального (амниотического) давления. Превышение величины базального амниотического давления в фазе сокращения матки на каждые +10 мм рт. ст. приводит к снижению интенсивности кровотока в интервиллёзном пространстве на 25 %. После достижения высоты давления в амниотической полости, превышающего 50 мм рт. ст., маточно-плацентарный кровоток практически прекращается.
- В миометральном контуре кровообращения (миометрий тела матки) интенсивность кровотока не зависит от величины внутримиометрального давления и остаётся постоянной на всём протяжении сократительного цикла. Природа изоволюмической регуляции интенсивности кровотока в этом контуре так до сих пор и не установлена.
- Цервикальный контур кровообращения в рожающей матке человека имеет две важных особенности. Во-первых, по мере повышения амниотического давления возрастает приток артериальной крови, который на пике эффективной схватки в полтора раза выше, чем в паузе. На интенсивность оттока крови из венозных депо шейки оказывает влияние давление, которое генерирует при прямом воздействии на ткань шейки плодный пузырь или предлежащая часть плода (конкретно – на постепенно вовлекаемую в оболочку плодовместилища часть укорачивающейся шейки). После сглаживания шейки интенсивность кровотока через венозные депо уже контролируется не только интенсивностью артериального притока, но и величиной давления на её ткани плодного пузыря или предлежащей части. При этом абсолютная величина давления, например, предлежащей части плода на ткани шейки во время родового сокращения в большей степени зависит не столько от величины давления в полости тела матки или её выходного сегмента, сколько от величины прироста объема полости этого сегмента. Сброс крови из шеечного контура идёт в венозное депо оболочки нижнего сегмента матки.
- Венозный дренаж всех контуров кровообращения формально функционирует в два этапа. Первый – кровь попадает вначале в венозные депо миометрия и затем из депо по выводным венам сбрасывается в венозные сплетения таза. Выводящие вены, дренирующие венозные депо тела матки, «перфорируют» наружный мышечный слой, а выводящие кровь из депо выходного отдела матки прямо анастомозируют с венами малого таза, не «перфорируя» при этом структуры контракционного кольца и фиксирующих матку связок. Но в обоих случаях сброс крови в венозные депо и из них в выводные вены и затем в венозные сплетения малого таза, контролируются внутримиометральным давлением. При этом точно установлено, что чем выше внутримиометральное давление, тем меньше объём эксфузии крови как в венозные депо миометрия, так и из них за пределы органа. При величине внутримиометрального давления, равного ≈ 30 мм рт. ст. эксфузия крови за пределы органа практически прекращается или достигает минимального уровня.
- Таким образом, во время фазного родового сокращения матки в гемодинамической системе органа могут возникать ситуации, при которых, в связи с возрастанием дефицита венозного дренажа или его полной блокады, в венозные депо миометрия и шейки матки кровь будет продолжать закачиваться. Возникает феномен силового депонирования крови.
- Силовое депонирование крови может оказывать влияние на величину функционального объёма венозных лакун, как в миометральном, так и в шеечном контурах кровообращения.
О фундаментальных механических свойствах миометрия матки человека
- Каждый утеромиоцит, независимо от места его локализации в функциональном сократительном модуле, слое или отделе миометрия обладает генетически детерминированным свойством развития спонтанной фазной ауторитмики.
- Базовым активатором этой спонтанной фазной ауторитмики является степень растяжения клетки. Утеромиоцит миометрия матки человека сокращается тогда и только тогда, когда он находится в оптимальной степени растяжения.
- В рожающей матке все сократительные элементы мышечных оболочек каждой из полостей при физиологически протекающей схватке находятся в степени растяжения, равной 2L0, где L0 – это длина сократительного элемента после полного опорожнения полости матки.
- Изменение длины оптимально растянутого сократительного элемента мышечной оболочки рожающей матки сопряжено с изменением радиуса кривизны элемента и изменением прямого расстояния между точками его фиксации и может происходить как в фазе сокращения, так и в фазе расслабления элемента только при соответствующих девиациях величины объёма гидравлического содержимого функциональных полостей органа.
- Растянутые утеромиоциты мышечной оболочки рожающей матки человека способны реализовать три основных типа фазного сокращения. От начала фазного сокращения и до его окончания (пика) происходит постоянное увеличение силы сокращения. Этот тип сокращения возможен только при сохранении длины растянутого утеромиоцита на всём протяжении фазы сокращения (изометрический режим). Сокращение утеромиоцита может происходить с прогрессирующим уменьшением его длины, достигающей минимальных значений на пике сокращения. Уменьшение длины утеромиоцита происходит при постоянной величине силы сокращений (изотонический режим). Растянутый утеромиоцит может уменьшать свою длину и при постоянно снижающейся величине линейного растяжения его миофибриллярного аппарата, что наблюдается при фазе расслабления и снижения нагрузки, ограничивающей степень растяжения клетки (ауксотонический режим).
- В каждом растянутом утеромиоците мышечной оболочки рожающей матки человека функционирует механорецепторный механизм обратной связи по растяжению, с помощью которого клетка не только воспринимает любое воздействие механической природы направленное на насильственное изменение длины растянутого миоцита, но и мгновенно и абсолютно адекватно реагирует на это влияние изменением натяжения своего миофибриллярного аппарата. С помощью прямой механической связи, независимо от её природы (морфологической, гидродинамической), существует способность мгновенно, без искажений и потерь, транслировать любую механическую информацию от одного сократительного элемента к другому или одновременно ко всем сократительным элементом конкретной мышечной оболочки. Механорецепторный механизм обратной связи по растяжению обеспечивает функциональную гомогенность и абсолютную синхронность морфофункционального состояния всех, объединённых прямой механической связью, сократительных элементов мышечной оболочки каждого функционального отдела матки.
Функциональная метрология рожающей матки человека
К началу первого периода родов в рожающей матке человека чётко выделяются три морфофункциональных отдела – тело матки, её нижний сегмент и шейка. Каждый из этих отделов имеет свои характерные метрологические параметры: форму, объём полостей, их площадь, толщину стенок (рис. 1, А и Б).
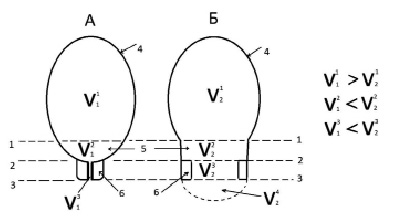
Рис. 1. Изменения формы и объёма полостей тела, нижнего сегмента и шейки матки в первом периоде родов. Обозначения: 1 – плоскость контракционного кольца; 2 – плоскость истмико-цервикального соустья; 3 – плоскость наружного зева шейки матки; 4 – тело матки; 5 – нижний сегмент; 6 – шейка матки. V11, V12,V13 – объёмы тела, нижнего сегмента и цервикального канала до начала первого периода родов; V21, V22,V23 – те же объёмы после завершения периода раскрытия; V24 – объём предлежащей части плода, вытесненный за пределы плоскости наружного зева шейки матки
К концу первого периода родов в каждом из функциональных отделов рожающей матки происходят изменения их метрологических характеристик, которые являются демонстративными маркерами сути биомеханических процессов, протекающих в каждом функциональном отделе органа. Так, к началу родов тело матки представлено образованием овоидной формы, с объемом V11, нижний сегмент – образованием в форме шарового сегмента с объемом V12 и шейка матки – образованием в виде толстостенного полого цилиндра с объемом цервикального канала равным V13. В начале первого периода родов шейка матки ещё не входит в состав оболочки плодовместилища. Нижний полюс плодного мешка со своим содержимым (передние воды, часть предлежащей части) локализуется в полости нижнего сегмента матки. К концу первого периода родов тело матки сохраняет свою форму и уменьшает объём своей полости. Следовательно, основная биомеханическая задача, которую решает в первом периоде родов сократительная система тела матки, состоит в силовом вытеснении частей объёма плодного мешка за пределы плоскости корпорально-истмического соустья, то есть за пределы контракционного кольца. За это же время нижний сегмент из шарового сегмента превращается в полый цилиндр с дистальным диаметром, равным проксимальному. Объём полости нижнего сегмента V12 возрастает примерно на 220 % (с ≈ 250 см3 до ≈ 550 см3). Одновременно с адаптивной трансформацией нижнего сегмента происходит сложный процесс двукратного переформатирования тканевых структур шейки матки. На первом этапе периода раскрытия шейка матки из полого цилиндра, благодаря процессам укорочения и сглаживания, превращается в шаровой слой, оболочка которого уже входит в состав дистального отдела плодовместилища.
На следующем этапе первого периода родов шейка матки из шарового слоя трансформируется в полый цилиндр, внутренний и наружный диаметры которого сравнимы с величиной диаметра цилиндра, в который превращается нижний сегмент матки. Полость цервикального канала с ≈ 14 см3 возросла до ≈ 200 см3 (+1650 %). При этом при физиологическом течении родов объём полости выходного отдела матки, уже состоящий из суммы полостей нижнего сегмента и шейки матки, возрастает, благодаря радиальному растяжению деформирующихся тканей. При этом длина стенки нижнего сегмента и шейки матки остаётся постоянной. Очень важным обстоятельством является то, что при втором этапе трансформации шейки матки из шарового слоя в полый цилиндр, одновременно с увеличением наружного диаметра зева, происходит вытеснение части объёма плодного мешка за пределы его плоскости, то есть в полость влагалища (объём V24), поскольку к моменту полного раскрытия наружного зева плодный пузырь вскрывается и передние воды в количестве ≈ 150 см3 изливаются, то величина объема V24 определяется частью объёма предлежащей части плода, вытесненной за пределы плоскости наружного зева (≈ 260 см3). То есть к моменту завершения первого периода родов в полости выходного отдела матки и за её пределами находится ≈ 1050 см3 объёма тела плода.
Следовательно, за весь первый период родов объём полости тела матки уменьшается на ≈1050 см3 (≈ – 23 %). При этом окружность тела матки уменьшается всего на 7 % при сохранении длины и нижнего сегмента, и шейки матки. Другими словами, процесс трансформации матки в первом периоде родов из органа плодоношения в орган плодоизгнания и во времени и в пространстве совмещен с процессом осевого перемещения тела плода из полости тела матки в полость её деформируемого выходного отдела.
Уменьшение объёма полости тела матки, осевое перемещение части тела плода из полости тела матки в полость деформируемого выходного отдела матки, увеличение объёма выходного отдела матки, связанное с осевым перемещением в его полость части объема тела плода – это одновременные, взаимосвязанные и взаимозависимо протекающие во время физиологической родовой схватки биомеханические события.
Миометрально-гемодинамический генез родовых сил и механизм конвертации их энергии в процесс «дистракции шейки матки»
При наличии в рожающей матке человека корпорально-истмического соустья (зона контракционного кольца), которая «тампонирована» телом плода, теоретически осевое перемещение его из одной функциональной полости в другую возможно только в одном случае, а именно, если экспульсионная сила, с помощью которой и может быть осуществлено это перемещение, будет больше, чем сила сопротивления деформируемых тканей выходного отдела матки. Эта сила формально противодействует осевому перемещению тела плода. Судя по тому, что деформирующая сила постоянно возрастает в каждом деформационном цикле, сопряженным с возрастанием степени напряжения мышечной оболочки тела матки в фазе её родового сокращения, сопротивление деформируемых тканей выходного отдела матки также растет по мере приближения цикла деформации к его пику. Чтобы успешно преодолевать возрастающее сопротивление деформируемых тканевых структур, необходимо, чтобы деформирующая ткани сила постоянно возрастала и достигала своего максимума к пику деформационного цикла.
Мы точно знаем, что мышечная оболочка тела матки и её нижнего сегмента не имеют между собой прямой механической связи. Следовательно, сила натяжения миофибрилл, повышающаяся во время схватки в клетках, локализованных в мышечной оболочке тела матки, не транслируется на клетки, локализованные в мышечной оболочке нижнего сегмента. При этом мы также точно знаем, что сила линейного натяжения утеромиоцитов в теле матки во время фазного родового сокращения органа постоянно нарастает от некоего минимума (базальный тонус) до некоего максимума (пик сокращения). Так, например, тангенциальное напряжение мышечной оболочки тела матки в паузе между схватками при давлении в полости тела равным ≈ 10 мм. рт. ст. составляет ≈ 84 г/см2, а при давлении в 45 мм рт. ст. на пике схватки ≈ 382 г/см2.
Поскольку внутриполостное давление в таких гидравлических системах, как тело рожающей матки, является функцией тангенциального напряжения его мышечной оболочки, то мы сегодня можем ещё раз утверждать, что мышечная оболочка тела матки в фазу родового сокращения органа сокращается в изометрическом режиме, то есть без укорочения длины растянутых утеромиоцитов. При этом, поскольку между мышечными оболочками тела и нижнего сегмента матки прямая механическая связь отсутствует, то вся энергия фазного сокращения мышечной оболочки тела матки конвертируется во внутриполостное давление. Таким образом, единственным фактором генерации силы, способной осуществлять осевое перемещение тела плода, является внутриамниотическое давление. Однако, согласно действию физических законов в закрытых гидравлических системах, подобных телу рожающей матки человека, силовое дискретное безвозвратное перемещение части объема тела плода за пределы полости тела матки обязательно должно сопровождаться адекватным уменьшением объема полости тела матки и укорочением длины растянутых сократительных элементов его оболочки. Автоматически происходящее укорочение длины растянутых сократительных элементов мышечной оболочки мгновенно переводит режим сокращения утеромиоцитов из изометрического в изотонический, что неизбежно приведёт к остановке генерации экспульсионной силы, осуществляющей осевое перемещение тела плода.
В рожающей матке человека в фазу сокращения органа документально доказанным фактом реальности перемещения части тела плода из одного функционального отдела матки в другой может служить как увеличение объема выходного отдела матки, так и увеличение диаметра плоскости наружного зева матки или диаметра плоскости истмико-цервикального соустья. Динамика изменения диаметра наружного зева – параметр, доступный для изучения и фиксации при проведении различных вариантов цервикодилатометрии, в том числе рутинных. Например, на рис. 2 отражены последовательно развивающиеся биомеханические события в матке у женщины при первых родах, которые по времени синхронизированы с изменением диаметра наружного зева сглаженной шейки матки в одну из ближайших после спонтанного излития околоплодных вод схваток при исходной величине диаметра наружного зева равного 43 мм.
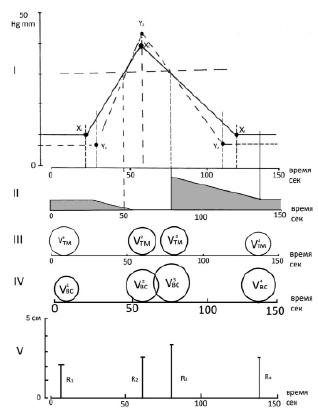
Рис. 2. Биомеханические события в матке у женщины при первых родах
Как видно на рис. 2, на всём протяжении фазы родового сокращения мышечная оболочка тела матки сокращается в изометрическом режиме (рис. 2, I). Объём полости тела матки (рис. 2, III) оставался постоянным (V1тм = V2тм), объём выходного отдела матки прогрессивно возрастал (V1нс < V2нс), и, соответственно этому, увеличивался радиус окружности наружного зева шейки (R1-R2). Увеличение объёма выходного отдела матки и увеличение диаметра наружного зева на пике сокращения (+Δ ≈ 2 мм) являются документальным подтверждением того, что эксцесс с перемещением части тела плода состоялся и состоялся он при сохранении величины объема полости тела матки. Если сопоставить динамику всех перечисленных выше биомеханических событий с динамикой венозного дренажа матки, то станет очевидным, что уже в начальной фазе сокращения миометрия тела матки прогрессивно возрастает дефицит венозного дренажа, что сопровождается избыточным наполнением кровью венозных депо миометрия. После достижения величины давления в полости тела матки, близком к 30 мм рт. ст., венозный дренаж органа либо полностью блокируется, либо остаётся на некотором минимальном уровне. И дальнейшая часть сокращения происходит на фоне силового депонирования крови в эти венозные резервуары миометрия, что увеличивает их объем и, следовательно, компенсирует убыль объема полости плодного мешка, обусловленную осевым перемещением тела плода.
Таким образом, именно феномен силового депонирования крови в венозные депо миометрия во время родовой схватки одновременно:
- обеспечивает сохранение изометрического режима сокращения мышечной оболочки тела матки;
- сохраняет на всём протяжении фазы сокращения миометрия необходимое постоянство объема гидравлического содержимого полости тела матки;
- обеспечивает сохранение эффективности воздействия на тело плода экспульсионной силы и модулирует эффективность процесса осевого перемещения тела плода.
Следовательно, к концу очередной фазы родового сокращения матки одновременно фиксируются:
- максимум тангенциального напряжения миометрия;
- максимум подъёма внутриполосного давления;
- максимум величины экспульсионной силы;
- максимум перемещения тела плода;
- максимум сопротивления этому силовому перемещению тела плода со стороны деформируемых тканей выходного отдела матки при сохранении объёма полости тела матки на всём протяжении первой фазы родового сокращения матки;
- максимум объёма депонируемой в венозные депо миометрия крови.
На рис. 2 также отражены основные биомеханические события, которые происходили в фазу родового сокращения матки в её выходном отделе. В это время и дистальная часть нижнего сегмента и шейка матки, включенная в оболочку плодовместилища, уже имела форму шарового слоя. Мы полагаем, что именно это последнее обстоятельство – включение шейки матки в состав оболочки плодовместилища – являет собой начало активной фазы первого периода родов, поскольку значительно меняет биомеханику деформационного силового процесса, происходящего в выходном отделе органа. На рис. 2, I, видно, что к началу сокращения матки давление в полости нижнего сегмента на 2–3 мм рт. ст. ниже, чем в полости тела матки. Этот факт подтверждает то, что между мышечными оболочками тела матки и её нижнего сегмента прямой механической связи нет. Механически (гидравлически) эти полости объединены только через тело плода. Начало сокращения мышечной оболочки нижнего сегмента начинается через короткое время (2–4 с) после начала сокращения мышечной оболочки тела матки. Однако прирост величины тангенциального напряжения и величины давления в оболочке и полости нижнего сегмента происходят с большей скоростью, чем в теле матки. То есть на определенном отрезке фазы сокращения матки градиент тангенциального напряжения и давления в пользу полости тела матки сохраняется – до тех пор, пока давление в обеих полостях не сравняется по значению. Затем градиент давлений инверсируется и, до пика сокращения, который оба функциональных отдела матки достигают одновременно, давление в полости нижнего сегмента становится и остается более высоким, чем в полости тела матки. Это означает, что реализация индивидуальной программы фазного сокращения утеромиоцитов, локализованных в оболочке нижнего сегмента, индуцируется импульсом дорастяжения, который генерируется в полости тела матки и из-за большой инерционности массы плода начинает транслироваться на оболочку нижнего сегмента с некоторой минимальной задержкой. Это также означает, что вся программа фазной активности оболочки нижнего сегмента от начала и до конца систолы схватки работает под перманентным воздействием импульса дорастяжения, генератором которого является осевое перемещение тела плода, при этом нижнему полюсу плодного мешка придаются свойства «гидравлического клина». И только при эффективной физиологической родовой схватке в конце фазы сокращения наблюдаются изменения величины тангенциального напряжения тканей нижнего сегмента и градиента давлений в полостях в пользу нижнего сегмента. Это является прямым доказательством того, что энергия силы импульса дорастяжения мышечной оболочки нижнего сегмента, генерируемая осевым перемещением плода, суммируется с энергией экспульсионной силы, генерируемой миометрально-гемодинамическим механизмом самого выходного отдела матки.
Этот «местный» источник генерации импульсов дорастяжения, как и в теле матки, являет собой мощное венозное депо шейки матки, которое, в отличие от миометрального, имеет артериально-венозные шунты и не имеет оболочки, способной активно менять величину внутриполостного давления в лакунарно трансформированных венах этого депо. Величина внутриполостного давления в сосудах депо, равно как их объем, прямо зависят от интенсивности артериального притока (на пике схватки он возрастает примерно в 1,5 раза), величины пассивного сопротивления к деформации тканей самой шейки и величины внешнего давления на ткани шейки элементов нижнего полюса плодного мешка, и возможности сбрасывания «излишков» крови в сосудистое депо оболочки нижнего сегмента. Все эти факторы организуют своё биомеханически значимое взаимодействие сразу же с началом процесса осевого перемещения плода. Силовое депонирование крови в венозные депо шейки обеспечивает силовой сброс крови в венозные депо нижнего сегмента. При этом генерируются прогрессивно нарастающие по мощности импульсы дорастяжения, которые на первой фазе систолы схватки способствуют увеличению скорости нарастания в оболочке нижнего сегмента величины тангенциального напряжения, что, в свою очередь, ускоряет рост давления в полости нижнего сегмента. А во второй фазе систолы, после инверсии градиента давлений в пользу полости нижнего сегмента, этот механизм становится дополнительным источником повышения экспульсионной силы, «помогающим», таким образом, телу матки преодолевать растущее сопротивление тканей шейки к деформации.
То есть во время физиологической родовой схватки в рожающей матке человека фактически функционируют два миометрально-гемодинамических механизма. Один функционирует в теле матки, второй – в выходном отделе органа. При этом первый механизм, функционирующий в теле матки, использующий энергию фазного сокращения мышечной оболочки и энергию силового депонирования крови в венозные миометральные депо, по существу, решает одну задачу – осевое перемещение части объёма концепта из полости тела матки в полость её выходного отдела. Второй миометрально-гемодинамический механизм, действующий в выходном отделе матки, решает несколько важнейших для нормального исхода родов биомеханически значимых задач.
Итак, как мы уже говорили выше, первая задача – усиление эффекта силы, генерируемой в полости тела матки, что увеличивает конкретную эффективность конкретного цикла осевого перемещения плода и, следовательно, увеличивает и эффект воздействия нижнего полюса плодного мешка (гидравлический клин) на деформирующиеся ткани шейки матки. Второй эффект силового депонирования крови в венозные депо самой шейки, циклы которого происходят на всём протяжении первого периода родов, связан с воздействием повышенного внутрисосудистого давления на ткани самой шейки матки, сопряженного с возрастанием объема огромного количества внутришеечных тонкостенных венозных лакун. При каждом цикле депонирования жидкая часть крови по градиенту давления из полости венозных лакун инфузируется в окружающие их ткани шейки. Ткани циклично «дообводняются» и столь же циклично контактируют с разного рода биологически активными веществами, которые меняют модуль упругости ткани, делая их более податливым для воздействия деформирующей силы. То есть по мере возрастания степени радиального растяжения тканей шейки их модуль упругости не только не возрастает, а при физиологически текущих родах, наоборот, имеет тенденцию к снижению.
Более драматичными для их адекватного понимания оказались биомеханические события, происходящие в рожающей матке человека в фазе расслабления (диастоле) родовой физиологической эффективной схватки. В момент достижения пика фазы сокращения оба функциональных отдела рожающей матки – и тело, и её нижний сегмент оказываются в принципиально различном морфофункциональном состоянии. Если объём и площадь полости тела матки оказались в состоянии, не отличавшемся от исходного, бывшего до начала схватки (V1TM = V2TM), то объём и площадь выходного отдела матки оказались увеличенными (V1НС < V2НС), что подтверждало и увеличение диаметра наружного зева шейки. Увеличение всех параметров, характеризующих морфофункциональное состояние выходного отдела матки, было прямо связано с перемещением в его полость части объёма концепта. Расслабление мышечных оболочек тела матки и нижнего сегмента происходит одномоментно, но сам процесс расслабления этих отделов происходит с разной скоростью. Скорость расслабления оболочки нижнего сегмента происходит с более высокими показателями, чем в теле матки. Градиент давления в пользу полости нижнего сегмента матки, имевшийся до начала расслабления матки, к моменту достижения давления в обеих полостях ≈ 30 мм рт. ст. нивелируется, и далее происходит инверсия давлений в пользу тела матки. Давление базового уровня в полости нижнего сегмента достигается на 8–10 с ранее, чем в полости тела матки. Именно с момента нивелирования градиента давления в полостях начинается буквально «обвальный» интенсивный отток венозной крови от органа, до этого момента «хранившийся» в гигантских венозных депо тела и шейки матки. Снижение интенсивности венозного оттока из депо тела матки в фазе расслабления органа достигает своего оптимума (базальный уровень дренажа до схватки) спустя ≈ 10–14 с после завершения фазы расслабления матки. К этому моменту объём полости тела матки уменьшается (V4ТМ) в результате изотонического сокращения функциональных сократительных модулей тела матки, строго адекватного уменьшению объёма полости тела. В это же самое время объём полости выходного отдела матки оказывается большим, чем исходный (V1нс< V4нс), что сопровождается увеличением диаметра растянутого наружного зева шейки. Причём конечный объём выходного отдела матки оказывается приблизительно равным объёму, который был достигнут на пике схватки (V2нс ≈ V4нс).
Увеличение диаметра наружного зева в итоге схватки также оказалось сопоставимым с величиной диаметра зева, достигнутого на пике сокращения матки. Как видно на рис. 2, IV, V, основные, в некотором смысле парадоксальные, биомеханические события в фазе расслабления матки происходят в выходном отделе матки в самом начале фазы диастолы (≈ 22 с) от пика схватки до начала венозного дренажа. В этот промежуток времени, казалось бы, при полном «биомеханической нейтралитете» со стороны тела матки (отсутствие динамики изменения объема полости тела матки, отсутствие признаков продолжающегося осевого перемещения тела плода) наблюдается интенсивное увеличение объема выходного отдела матки и возрастание диаметра растянутого зева матки (V2НС < V3НС, R2 < R3). Поскольку увеличение объёма выходного отдела матки и увеличение диаметра наружного зева происходило на фоне расслабления мышечной оболочки нижнего сегмента, и затем, на фоне этого же, далее продолжающегося расслабления, эти же оболочки уменьшили свои параметры до их финальных, полученных в итоге схватки значений, то можно полагать, что девиации этих показателей не были прямо обусловлены участием в их генезе изменениями морфофункционального состояния мышечных элементов оболочки нижнего сегмента матки. Согласно биомеханической логике деформационно-силовых процессов, происходящих в выходном отделе рожающей матки человека во время фазы сокращения органа, достигнутая на пике схватки степень дорастяжения диаметра наружного зева немедленно и необратимо фиксируется внедрением в него нижнего полюса плодного мешка. Более того, по мере увеличения диаметра наружного зева матки внедряющаяся в него дистальная часть концепта, перемещается и за пределы плоскости наружного зева матки. Парадоксально, но, несмотря на значительное по масштабам увеличение диаметра наружного зева матки, происходящее на протяжении этой фазы диастолы схватки, которое достигает своего максимума на момент обратной инверсии градиента давления в функциональных полостях матки и начала венозного дренажа органа, диаметр дорастянутого зева по мере дальнейшего расслабления матки начинает уменьшаться и достигает своих конечных, значительно меньших, итоговых размеров. То есть, поскольку фиксации максимального размера дорастянутого диаметра наружного зева матки на этом этапе схватки не происходит, есть все основания полагать, что сам процесс его дилатации в эту фазу родовой схватки прямо не связан ни с генерацией экспульсионной силы, ни с осевым перемещением тела плода. И, поскольку процесс его обратной деформации прямо связан с началом и продолжением венозного дренажа органа, то есть все основания связать факт его дилатации в этот момент схватки с особенностями гемодинамики самой шейки матки. В связи с изложенным возникают два вопроса. Первый: «Если увеличение объема выходного отдела матки и диаметра наружного зева в начальной фазе диастолы родовой схватки не оказывает никакого влияния на конечную эффективность конкретного сокращения органа, то для чего, с какой целью и именно только при эффективной физиологической родовой схватке генерируется этот своеобразный биомеханический парадокс?» Вопрос второй: «Если понятно, как и зачем феномен силового депонирования крови в фазу эффективного родового сокращения матки принимает прямое участие в процессах генеза родовых сил и осевого перемещения тела плода, то как этот же феномен способен влиять на рост объёма выходного отдела матки и увеличения диаметра наружного зева органа в фазу его расслабления, в частности в начальной фазе диастолы схватки?» Мы точно знаем, что процесс «расслабления – укорочения» растянутого миометрия может происходить только при уменьшении объёма полости тела матки. Мы так же точно знаем, что увеличение объёма полости тела матки даже в фазу расслабления мгновенно остановит фазу его расслабления и укорочения. Нам известно, что до начала венозного дренажа органа на всём протяжении этой начальной фазы диастолы расслабление миометрия тела матки происходит без его укорочения, то есть в изометрическом режиме и, следовательно, при сохранении постоянства объёма полости тела матки. Если режим силового депонирования крови в сосудистые депо тела матки в эту фазу схватки сохраняется, объём полости тела матки при отсутствии осевого перемещения плода сохраняет свою исходную (до начала расслабления матки) величину, то подобного рода биомеханическая ситуация может быть реализована только в одном случае – если инфузируемый в венозное депо миометрия «избыточный» объем крови будет эксфузироваться из венозных депо миометрия в венозное депо нижнего сегмента и шейки матки. Поскольку в то же время процесс «внутреннего» депонирования крови в венозные депо шейки матки продолжается, то «наружная» и «внутренняя» система депонирования одновременно увеличивают объём полости нижнего сегмента матки и объем интрацервикального депо, что и приводит к эффекту «эректилизации» криволинейной формы оболочки шейки, к её некоторому спрямлению и, естественно, растяжению наружного зева. При этом, и это очень важно, увеличение объема нижнего сегмента матки не сопровождается увеличением длины уже оптимально растянутых функциональных сократительных модулей её оболочки. Радиальное растяжение соединительнотканных структур зоны истмико-цервикального соустья «раздвигает» зоны фиксации нижних оконечностей модулей, увеличивая расстояние между ними, что неизбежно оказывает влияние на модуль упругости вязко-эластической структуры этой области шейки матки. А феномен «эректилизации» тканей шейки и спрямления её оболочки, связанный с силовым увеличением объёмов её венозных депо, сопровождается точно таким же действием на тканевые структуры отдельных участков тканей шейки по мере того, как в процессе её укорочения они будут входить с состав оболочки плодовместилища.
Есть основания полагать, что физиологический смысл этого уникального биомеханического события, происходящего в начальной фазе диастолы родовой схватки, заключается в том, чтобы с помощью биофизического и биохимического воздействия на тканевые структуры шейки снизить модуль их упругости и «подготовить» шейку матки к следующему сократительному циклу, повышая, таким образом, эффективность каждой последующей родовой схватки.
Итак, мы представили краткое содержание нашей авторской версии миометрально-гемодинамической концепции биомеханики физиологической родовой схватки. С более подробным описанием концепции, равно как и с её научной компетентностью и доказательной легитимностью, можно познакомиться в монографиях и статьях, имеющих фундаментальное значение [12, 15, 17].
Изложенная выше авторская версия миометрально-гемодинамической концепции биомеханики физиологической родовой схватки не только раскрывает сущность биофизических механизмов генерации родовых сил и механизмов конвертации их энергии в полезную внешнюю работу по дилатации шейки матки, что имеет огромное значение для практики акушерства родильного зала, но и позволяет понять значение необходимости поддержания оптимальной величины базального тонуса матки и готовности сосудистой системы шейки матки к родам, как основных гарантов эффективного и физиологического течения родового процесса. Наши многочисленные исследования подтверждают, что игнорирование врачом, ведущим роды, этих двух постулатов гарантированно обеспечивает их осложнённое течение, что часто, даже при проводимой стандартной коррекции, приводит к неблагоприятному воздействию на мать и плод.
Библиографическая ссылка
Савицкий А.Г., Савицкий Г.А. БИОМЕХАНИКА ФИЗИОЛОГИЧЕСКОЙ РОДОВОЙ СХВАТКИ (МИОМЕТРАЛЬНО-ГЕМОДИНАМИЧЕСКАЯ КОНЦЕПЦИЯ) // Научное обозрение. Медицинские науки. 2021. № 6. С. 41-53;URL: https://science-medicine.ru/en/article/view?id=1216 (дата обращения: 18.07.2026).
DOI: https://doi.org/10.17513/srms.1216