 science-review.ru
science-review.ru
Scientific journal
Научное обозрение. Медицинские науки
ISSN 2500-0780
ПИ №ФС77-57452
EFFICACY OF ADEMETIONINE AND URACIL-DERIVED COMPOUND IN CORRECTING CHANGES IN THE EXPRESSION OF ANTIOXIDANT DEFENSE GENES IN THE KIDNEYS OF RATS UNDER CHRONIC THIOACETAMIDE EXPOSURE

Введение
Тиоацетамид (ТАА) представляет собой сероорганическое соединение, которое не встречается в природе, является искусственно созданным веществом и активно применяется в различных сферах человеческой деятельности. ТАА широко используется в качественном неорганическом анализе в качестве источника сульфид-ионов, что позволяет получать сульфиды металлов при взаимодействии с водными растворами таких катионов металлов, как никель, свинец и ртуть. Его применяют в качестве стабилизатора моторного топлива и ускорителя вулканизации каучука, в кожевенной, текстильной и бумажной промышленности. ТАА ранее также применялся в качестве фунгицида, в настоящее время широко используется в экспериментальной токсикологии как модельное соединение, способное вызывать повреждения печени и других органов [1]. Токсичность ТАА обусловлена его метаболическим превращением в печени с участием цитохрома P450 до высокореактивных метаболитов – тиоацетамид-S-оксида и тиоацетамид-S-диоксида. Эти метаболиты проявляют цитотоксическое действие как за счет ковалентного связывания с макромолекулами, так и посредством индуцированного оксидативного стресса, сопровождающегося генерацией свободных радикалов и истощением антиоксидантных систем защиты. Реактивные метаболиты тиоацетамида оказывают токсическое действие посредством двух основных механизмов: прямого взаимодействия с клеточными макромолекулами (образуя ковалентные аддукты с белками, липидами и нуклеиновыми кислотами) и опосредованного воздействия через индукцию оксидативного стресса. Последний сопровождается массивной генерацией свободных радикалов, истощением антиоксидантных систем [2]. При этом даже однократное воздействие ТАА способно инициировать повреждение печени и нарушение ее детоксикационной функции, что влечет за собой системное распространение токсических соединений, включая аммиак, обладающий нефротоксичностью [3]. Кроме того, кинетика образования и накопления метаболитов ТАА характеризуется насыщаемостью и значительной индивидуальной вариабельностью, что затрудняет прогнозирование степени и направленности органных повреждений [4].
Почки, играя ключевую роль в элиминации метаболитов и поддержании гомеостаза, становятся уязвимыми мишенями при нарушении функции печени и системной оксидативной нагрузке. Острые и хронические поражения почек, развивающиеся на этом фоне, ассоциированы с высоким риском инвалидизации и летального исхода [5, 6]. В настоящем исследовании авторы сосредоточились на оценке потенциальных терапевтических стратегий, направленных на коррекцию почечных нарушений при остром воздействии модельного токсиканта. Одним из перспективных направлений является использование средств с антиоксидантными и мембраностабилизирующими свойствами, способных эффективно вмешиваться в патогенетические механизмы токсического действия на клеточном уровне. В частности, адеметионин (S-аденозилметионин) и комбинация 5-гидрокси-6-метилурацила с ацетилцистеином могут потенциально модулировать экспрессию генов антиоксидантной защиты и снижать выраженность тканевого окислительного стресса.
Цель исследования – оценить влияние адеметионина и комплекса 5-гидрокси-6-метилурацила с ацетилцистеином на экспрессию генов антиоксидантного ответа (Hmox1, Nqo1) в почечной ткани крыс при хроническом воздействии тиоацетамида
Материалы и методы исследования
Исследование было спланировано с учетом международных рекомендаций ARRIVE по гуманному использованию лабораторных животных. Протокол эксперимента утвержден локальным биоэтическим комитетом (решение № 01-02 от 08.02.2024) и полностью соответствовал нормативным требованиям РФ к работе с экспериментальными животными.
56 аутбредных крыс-самцов массой 170–190 г (возраст 12–14 недель) содержались в стандартных условиях вивария. Температурный режим поддерживался на уровне 21–26 °C, животные содержались при 12-часовом световом цикле с автоматическим переключением. Был обеспечен свободный доступ к воде и стандартному корму для лабораторных животных. Перед началом эксперимента животные прошли 5-дневную акклиматизацию.
Животные были рандомизированы на четыре группы (n = 14 для каждой группы). Первая группа являлась положительным контролем и получала раствор тиоацетамида (ТАА) в дозе 50 мг/кг массы тела. Вторая и третья группы получали аналогичное токсическое воздействие на фоне коррекции – адеметионина в виде лекарственного препарата «Самеликс», (ООО Фирма «Фермент», Россия) в разовой дозе 25 мг/кг массы тела за 1 ч до ТАА (ТАА + С); либо экспериментального комплексного соединения 5-гидрокси-6-метилурацила с ацетилцистеином в разовой дозе 500 мг/кг массы тела, разработанного Уфимским институтом химии Российской академии наук [7]. Четвертая группа являлась отрицательным контролем и получала эквивалентный объем физиологического раствора (К). Продолжительность эксперимента составила 100 дней с промежуточным контролем на 50-й день.
По окончании эксперимента животных подвергали эвтаназии путем декапитации. Почечную ткань забирали немедленно после эвтаназии для последующего молекулярно-генетического анализа. РНК выделяли из тканевых образцов органа с использованием набора ExtractRNA («Евроген», Россия) в строгом соответствии с инструкциями производителя. Синтез кДНК осуществляли с применением набора реактивов MMLV RT kit и праймеров олиго(dT)15 («Евроген», Россия). Для исследования транскрипции генов использовали метод полимеразной цепной реакции в режиме реального времени, при котором применяли специфичные олигонуклеотидные праймеры и интеркалирующий краситель SYBR Green. Ген Gapdh служил внутренним контролем. Для оценки относительной экспрессии генов использовался метод, предложенный K.J. Livak и T.D. Schmittgen [8]. Для определения разницы между группами рассчитывался показатель ΔΔCT, который представлял собой разность между средним значением ΔCT группы сравнения и группы контроля. Результирующее значение, полученное посредством экспоненциального преобразования этой разности, интерпретировалось как «fold change» (FC), что позволяло оценить, насколько изменена экспрессия целевого гена в исследуемой группе относительно контроля. Были выбраны ген, кодирующий гем-оксигеназа 1 (Hmox1), который считается одним из наиболее чувствительных и надежных индикаторов клеточного окислительного стресса (идентификатор гена согласно NCBI: 24451) [9]. Изучали уровень экспрессии гена. кодирующего NAD(P)H-хиноноксидоредуктазу 1 (Nqo1) – ключевой фермент детоксикации, играет важную роль в хемопротекции, катализирующий восстановление хинонов и участвующий в антиоксидантной защите посредством генерации восстановленных форм убихинона и витамина Е (идентификатор гена согласно NCBI: 24314) [10].
Статистический анализ проводили с использованием программного обеспечения SPSS Statistics 21.0 (IBM, USA). В качестве критерия нормальности распределения признаков в изучаемых группах использовали критерий Колмогорова – Смирнова. При нормальном распределении данных для оценки значимости различий между группами использовали однофакторный дисперсионный анализ (ANOVA) и апостериорные критерии Тьюки и Тамхейна. Данные представлены как среднее арифметическое и стандартная ошибка. Статистический анализ данных проводился с использованием метода бутстрап (Bootstrap), что обеспечивало надежность и воспроизводимость результатов через многократную генерацию выборок методом Монте-Карло. Различия считали статистически значимыми при р < 0,05.
Результаты исследования и их обсуждение
В отличие от хорошо изученных гепатотоксических эффектов, нефротоксическое действие ТАА остается менее исследованным. При длительном воздействии описаны гломерулярные нарушения, застойные явления, повышение уровней креатинина и мочевины, а также активация провоспалительных и оксидативных путей [11, 12]. Однако молекулярные механизмы этих изменений, особенно в условиях хронической нагрузки, остаются недостаточно ясными, что обусловливает актуальность оценки экспрессии антиоксидантных генов в почечной ткани.
Выбор генов Hmox1 и Nqo1 в качестве маркеров клеточного ответа обусловлен их ключевой ролью в системе антиоксидантной защиты. Hmox1 кодирует гемоксигеназу-1 – чувствительный индикатор оксидативного стресса, Nqo1 – фермент второй фазы детоксикации, участвующий в восстановлении хинонов и защите от реактивных метаболитов [9, 10]. Оба гена используются как универсальные маркеры включения клеточной защиты при воздействии ксенобиотиков.
Полученные в проведенном исследовании результаты демонстрируют ограниченную реактивность антиоксидантных генов в почках на фоне хронического воздействия ТАА. Как видно на рис. 1, на 50-й день воздействия кратность экспрессии гена Nqo1 в ткани почек крыс статистически значимо не отличалась между контрольной группой и группой ТАА (–7,26 ± 0,28 против –7,17 ± 0,09; p = 0,783). Однако при сопоставлении группы ТАА с группой ТАА + Самеликс было зафиксировано достоверное повышение экспрессии Nqo1 (–7,17 ± 0,09 против –6,82 ± 0,09; p = 0,047), что согласуется с известным эффектом адеметионина на метаболизм глутатиона и метилирование промоторов антиоксидантных генов [13, 14]. В группе ТАА +М уровень экспрессии не отличался от контроля и от ТАА (p > 0,05), что может быть связано с реализацией эффекта комплекса 5-гидрокси-6-метилурацила с ацетилцистеином в обход транскрипционной активации Nrf2, за счет прямого донорства SH-групп или непрямого модулирования репаративных путей [15]. На 100-й день воздействия статистически значимых изменений экспрессии Nqo1 ни в одной из групп по сравнению с контролем выявлено не было (p > 0,05).
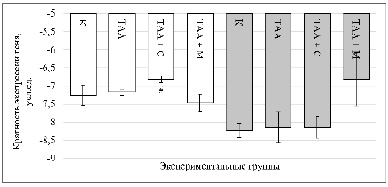
Рис. 1. Изменение кратности экспрессии гена Nqo1 в ткани почек крыс при субхроническом (50 дней) и хроническом (100 дней) воздействии ТАА и на фоне фармакологической коррекции. По оси ординат – относительная кратность экспрессии (нормализована к Gapdh), по оси абсцисс – экспериментальные группы. * – статистически значимые различия (p < 0,05) по сравнению с группой отрицательного контроля (К) соответствующего временного периода; # – статистически значимые различия (p < 0,05) по сравнению с группой ТАА соответствующего временного периода. Белые столбцы – 50 дней воздействия, серые – 100 дней Источник: составлено авторами на основании полученных экспериментальных данных
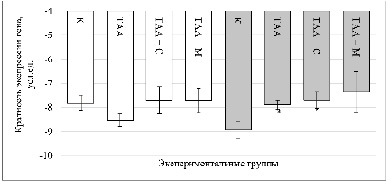
Рис. 2. Изменение кратности экспрессии гена Hmox1 в ткани почек крыс при субхроническом (50 дней) и хроническом (100 дней) воздействии ТАА и на фоне фармакологической коррекции. По оси ординат – относительная кратность экспрессии (нормализована к Gapdh), по оси абсцисс – экспериментальные группы. * – статистически значимые различия (p < 0,05) по сравнению с группой отрицательного контроля (К) соответствующего временного периода; # – статистически значимые различия (p < 0,05) по сравнению с группой ТАА соответствующего временного периода. Белые столбцы – 50 дней воздействия, серые – 100 дней Источник: составлено авторами на основании полученных экспериментальных данных
Как видно на рис. 2, экспрессия гена Hmox1 на 50-й день достоверно не отличалась между всеми исследуемыми группами, что указывает на отсутствие выраженной активации антиоксидантного ответа в изученные сроки воздействия. К 100-му дню воздействия наблюдалось статистически значимое снижение экспрессии Hmox1 в группе ТАА по сравнению с контролем (–8,93 ± 0,32 против –7,89 ± 0,20; p = 0,036), что может отражать истощение компенсаторного ответа на фоне хронической нагрузки. При этом в группе ТАА + Самеликс также было зафиксировано достоверное различие по сравнению с контролем (–8,93 ± 0,32 против –7,69 ± 0,34; p = 0,042), тогда как в группе ТАА + МГ-10 изменения не достигали статистической значимости (p > 0,05).
Суммарно данные указывают на то, что молекулярный отклик почек при хроническом воздействии ТАА развивается медленно, с ограниченной включенностью транскрипционных антиоксидантных механизмов. Согласно результатам проведенного исследования, адеметионин оказывал более выраженное действие на экспрессию Nqo1 на раннем этапе, но не предотвращал снижение Hmox1 в поздний срок, тогда как комплекс 5-гидрокси-6-метилурацила с ацетилцистеином не модулировал Nqo1, но демонстрировал тенденцию к сохранению кратности экспрессии на уровне отрицательного контроля гена Hmox1 к 100-му дню. Это может свидетельствовать о различиях в точках приложения действия препаратов: адеметионин преимущественно активирует эпигенетические и метаболические пути антиоксидантного ответа, а комплекс 5-гидрокси-6-метилурацила с ацетилцистеином действует скорее через стабилизацию митохондрий и подавление вторичного воспалительного каскада, не вовлекая напрямую путь Nrf2.
Полученные данные важны для развития направлений, связанных с фармакологической коррекцией нефротоксических эффектов, возникающих при хроническом токсическом воздействии. Показанная ограниченность включения антиоксидантного ответа в ткани почек при экспозиции тиоацетамидом может указывать на латентное течение патологического процесса. Перспективным направлением также является расширение панели молекулярных маркеров с включением генов воспалительного ответа, апоптоза и митохондриального гомеостаза, а также комплексное сопоставление молекулярных данных с биохимическими и морфологическими характеристиками. Это позволит сформировать более полное представление о патогенезе ТАА-индуцированной нефротоксичности и создать обоснованную платформу для оценки эффективности коррекционных воздействий.
Заключение
В условиях хронического воздействия тиоацетамида в почечной ткани крыс не зафиксировано выраженной активации генов антиоксидантного ответа. Установлено, что адеметионин способен усиливать экспрессию Nqo1 на ранних сроках воздействия, тогда как 5-гидрокси-6-метилурацил с ацетилцистеином статистически значимых изменений не показал, что свидетельствует о различии в механизмах их действия.
Conflict of interest
Financing
Библиографическая ссылка
Рябова Ю.В., Смолянкин Д.А., Каримов Д.Д., Кудояров Э.Р., Якупова Т.Г., Репина Э.Ф., Каримов Д.О. ЭФФЕКТИВНОСТЬ АДЕМЕТИОНИНА И ПРОИЗВОДНОГО УРАЦИЛА В КОРРЕКЦИИ ИЗМЕНЕНИЙ КРАТНОСТИ ЭКСПРЕССИИ ГЕНОВ АНТИОКСИДАНТНОЙ ЗАЩИТЫ В ПОЧКАХ КРЫС ПРИ ДЛИТЕЛЬНОМ ВОЗДЕЙСТВИИ ТИОАЦЕТАМИДА // Научное обозрение. Медицинские науки. 2025. № 4. С. 12-16;URL: https://science-medicine.ru/en/article/view?id=1449 (дата обращения: 22.06.2026).
DOI: https://doi.org/10.17513/srms.1449